Настоящий документ может без ограничений распространяться по электронным сетям на некоммерческой основе при условии сохранения цельности текста, включая настоящее уведомление и указания имени автора.
Любое другое использование документа, включая размещение документа на серверах, может производиться лишь с согласия автора при соблюдении его прав.
Авторские права зарегистрированы.
E-mail: bigmax2@yandex,ru
НЕОБХОДИМОЕ ВВЕДЕНИЕ
Два человека должны разделить со мной "ответственность" за появление этой работы. Это мой брат С.С. Москвитин и его жена Н.С. Москвитина.
Сергей Степанович много лет работает директором Зоологического музея Томского государственного университета (ТГУ). Кроме этого он является председателем Томского отделения Мензбирского орнитологического общества АН РФ и председателем Томского отделения Союза охраны птиц России.
Нина Сергеевна заведует кафедрой биологии позвоночных и экологии в ТГУ. Она доктор биологических наук.
Они пробудили во мне и всячески поддерживали мой интерес к биологии и проблеме происхождения биологического вида, всегда находили ответы на мои вопросы и никогда не отказывались обсудить любые мои идеи, в том числе самые бредовые. Без них появление этой работы на свет было бы невозможно.
За все это и многое другое мой низкий им поклон.
Необходимо, однако, отметить, что ответственность, да еще если это слово взято в кавычки, не означает вину. В конце концов и у вполне приличных родителей бывают беспутные дети.
Мой успех или неудачу со мной разделит моя семья.
Ну а мне остается надеяться, что в конечном счете второй эпиграф по отношению к этой работе приобретет тот смысл, который я в него вложил.
А. Москвитин
ПРЕДИСЛОВИЕ К ПЯТОЙ РЕДАКЦИИ
Пятая редакция "Третьего постулата" существенно отличается от предыдущих четырех. Отличие состоит в первую очередь в том, что в данной редакции учтены некоторые результаты обсуждения работы в Интернете. Кроме того я счел необходимым изменить форму изложения материала - в конце концов "Третий постулат" это не диссертация, представленная на соискание научной степени и прежняя псевдонаучная форма изложения работе только вредила. Сейчас же работа представляет из себя именно то, чем она и является - изложение результатов инженерного подхода к проблеме видообразования.
Что привело к появлению этой работы? Во-первых, ставший в свое время для меня откровением факт, что учение Ч. Дарвина не является окончательным словом в проблеме возникновения новых видов. Во-вторых - интерес к биологии вообще, разбуженный и поддерживаемый теми, о ком я сказал во введении. Ну и в-третьих - своеобразная извращенность мышления, заставляющая меня постоянно сводить известные мне в какой-либо области факты в некое подобие системы, что в случае положительного итога дает мне ощущение психологического комфорта.
Немного об истории написания "Третьего постулата".
Первая редакция была написана в феврале 2001 года и размещена в Интернете в 2002 году. Редакции вторая, третья и четвертая базировались на первой и являлись ее развитием. Что из себя представляет пятая редакция, я уже сказал. Редакции 5.2 и 5.3 соотносятся с этой пятой редакцией так же как вторая, третья и четвертая с первой, то есть отличаются от пятой небольшими изменениями и дополнениями.
В настоящей редакции - редакции 5.4 - появилась пятнадцатая глава: "Геном, как физическая система" и сделаны некоторые дополнения в главе 14, а именно: добавлена схема возникновения и вымирания видов, принципиально отличающаяся от известного всем "эволюционного древа".
Нетрудно заметить, что содержание главы 15 находится в некотором конфликте с содержанием остальных глав, что касается в первую очередь соображений о "балластной части ДНК". Тем не менее, конфликт этот я решил оставить неразрешенным как для иллюстрации неоднозначности подхода к вопросу, так и потому, что истина, как водится, лежит где-то посередине.
МОСКВИТИН А.К.
ТРЕТИЙ ПОСТУЛАТ
К вопросу о происхождении видов
(Редакция 5.4)
Все можно наладить, если вертеть
в руках достаточно долго.
Второй закон Вышковского
Решение любой проблемы становится
очевидным после того, как оно найдено.
Автор
Глава 1
Является ли "Третий постулат" научной работой?
Прежде чем ответить на вопрос, вынесенный в заголовок этой главы, необходимо определить критерий научности. Мое личное мнение мало кому может показаться убедительным, так как оно принадлежит лицу заинтересованному, поэтому необходимо мнение постороннего.
Далее цитируется профессор математики университета в Квебеке (Канада) Уильям С. Хэтчер. Цитата взята из статьи "Размышления о Всевышнем", опубликованной в журнале TERMINATOR N2-3 за 1994 г. (Санкт-Петербург, "Комкон"). Статья представляет сокращенный вариант лекции, прочитанной профессором 11 марта 1994 г. в Доме ученых в Лесном в Санкт- Петербурге.
ПРИРОДА НАУЧНОГО ДОКАЗАТЕЛЬСТВА
Прежде всего следует принять к сведению, что так называемое научное доказательство объясняется неоднозначно и, как правило, ошибочно.
Научное доказательство истинности какого-либо явления или факта слагается из двух этапов: многократного наблюдения и регистрации измерительными приборам и явления или факта и абстрактного теоретизирования над собранными данными, завершающегося выдвижением гипотезы или теории.
Далее происходит проверка выдвинутых предположений путем более точных наблюдений и измерений, Однако в силу ограниченности чувственного аппарата и нервной системы человека-исследователя нельзя исключить ошибок в этих процессах. Особенно верно сказанное в отношении предельно малых (микроскопических) объектов и явлений и предельно больших и сверхудаленных звездных систем и квазизвездных образований. Так или иначе, следует подчеркнуть, что в целом истинность фактов или явлений в любых масштабах в принципе всегда относительна, и широко распространенное убеждение в безусловной истинности научно установленных фактов или явлений есть заблуждение.
Новые эмпирические выводы, дедуцированные из гипотезы или теории, могут быть подтверждены опытом, и в этом случае мы считаем их вполне научно обоснованными, а научный взгляд полагаем справедливым и имеющим силу. Однако это совершенно не означает, что в отдаленном будущем не откроются факты или явления, не вмещающиеся в выстроенную систему взглядов, либо решительно опровергающие ее. То есть мы вновь вынуждены говорить об относительности научного познания. Более того, чисто формально в математической логике доказано, что можно предложить бесконечное количество взаимно исключающих теорий, объясняющих данный набор фактов или явлений.
Из вышесказанного безусловно следует фундаментальное заключение: ни одна из научных истин не может считаться абсолютно доказанной, а представление о возможности абсолютного доказательства само по себе не является научным. Таким образом, широко распространенное убеждение, что главные свойства научного доказательства есть его абсолютность и точность (в противоположность "относительности" и "неточности" истины в религии), ведут к заблуждению.
Можно иначе сформулировать наш вывод: научно обоснованное объяснение нового факта или явления должно характеризоваться более высокой вероятностью или правдоподобностью, чем любые другие объяснения...
Мне нечего добавить к мнению профессора. Остается только приложить усилия к тому, чтобы мой взгляд на проблему видообразования отличался "более высокой вероятностью или правдоподобностью".
Глава 2
Мутации
Начать изложение своих взглядов на проблему происхождения видов логичнее всего с определения основных понятий, главным из которых в эволюционных теориях является понятие мутации.
Мутации (от лат.mutatio - изменение), внезапные, естественные или вызванные искусственно наследственные изменения генетич. материала, приводящие к изменению тех или иных признаков организма. (Биологический энциклопедический словарь)
Несмотря на авторитетность определения, прежде чем двигаться дальше стоит повнимательнее "осмотреться в отсеках", и в первую очередь необходимо детализировать основное понятие приведенного определения: изменение генетического материала.
Под генетическим материалом в дальнейшем предлагается понимать генно-хромосомный набор - геном - несущий в себе наследственную информацию о всех признаках, присущих данной биологической особи.
Такой подход позволяет разделить изменения генетического материала на изменения, происходящие в отдельных генах, изменения, происходящие в хромосомах и изменения, затрагивающие весь геном особи.
Мутации в настоящей работе рассматриваются исключительно в плане их наследуемости, то есть при условии возникновения мутаций в половых, а не в соматических клетках.
Изменения в отдельных генах, приводящие к искажению их исходной - домутационной - структуры могут быть вызваны как внутренними, так и внешними причинами.
К внутренним причинам относятся ошибки репликации ДНК при делении клеток - мейозе и митозе. Известно, что репликация ДНК - очень точный процесс, и частота ошибок при его протекании, приводящих к спонтанным точковым мутациям, составляет около 10 в степени минус 9 на нуклеотид на поколение. Тем не менее саму возможность мутаций генов за счет ошибок при репликации ДНК в дальнейшем будем иметь в виду.
Спонтанные мутации генов для простоты также будем считать вызванными внутренними причинами.
Внешними причинами изменения исходной структуры генов являются мутагенные факторы физической и химической природы. Мутагенные факторы либо увеличивают частоту спонтанных мутаций, либо непосредственно влияют на структуру генов.
Представляется достаточно сомнительным сам факт, что в результате мутаций генов происходит упорядоченное перестроение закодированной в них информации, наоборот, гораздо логичнее предположить, что мутации могут только исказить эту информацию, вызвать, если провести аналогию с энергией замкнутой физической системы, только возрастание энтропии информации, содержащейся в генах.
Справедливость такого предположения неоднократно подтверждена фактами. Если до возникновения индустрии радиоактивных материалов к числу мутагенных факторов относились космическое излучение, достигающее поверхности Земли, излучения природных месторождений радиоактивных материалов и природные химические мутагены, то с наступлением ядерной эры и с развитием химической промышленности количество мутагенных факторов и число случаев их воздействия на живые организмы возросло неимоверно. Тем не менее, результат таких воздействий один - потомство особей, подвергшихся воздействию мутагенных факторов несет на себе печать уродства, зачастую несовместимого с жизнью.
Примером мутационных изменений, происходящих в хромосомах может служить неравный кроссинговер, в результате которого происходит увеличение количества генов, контролирующих один и тот же признак. Неравный кроссинговер может вызвать изменение контролируемого продублированными генами признака, но не приводит к появлению новых признаков.
В результате мутационных изменений в геноме происходит изменение числа хромосом и их макроструктуры. Эти изменения выражаются либо в анэуплодии - утрате или добавлении отдельных хромосом, либо в полиплодии - добавлении целых гаплоидных наборов хромосом.
Изменение числа хромосом в геноме в некоторых случаях ведет к изменению влияния отдельных генов, причем связанные с этими генами признаки могут меняться значительно сильнее, чем при генных мутациях.
Рассматривая мутации в общем плане необходимо сказать, что ни один из факторов, их вызывающих, не привносит в наследственную ДНК "строительный материал" для образования нового гена, и, следовательно, действует в рамках неизменного объема генетической информации.
Справедливость высказанного суждения по отношению к внешним мутагенным факторам достаточно очевидна. Что же касается внутренних процессов в геноме, то здесь ситуация не так наглядна. Ведь, например, в результате неравного кроссинговера может произойти увеличение количества генов в геноме. Но внутренние процессы чаще всего лишь дублируют уже имеющуюся в геноме информацию. Дублирование же не есть привнесение информации новой. Аналогия: прочитав два экземпляра одного и того же учебника, вы не увеличите количества полученных знаний по сравнению с тем, как если бы ограничились чтением (естественно, внимательным) только одного экземпляра.
В эволюционных теориях мутациям отводится главенствующая роль. Именно они считаются фактором, приводящим к возникновению у биологических особей новых, ранее отсутствовавших у них признаков.
Поскольку явление изменения числа генов в хромосомах и хромосом в геноме и последствия этих изменений достаточно хорошо известны, очевидно, что они не являют собой тот магистральный путь, на котором происходит возникновение новых видов. Поэтому в дальнейшем мы будем говорить о роли в процессе видообразования преимущественно генных мутаций.
Формально к мутациям может быть отнесена генетическая рекомбинация - обмен генетическим материалом, происходящий при размножении бактерий. Принципиальное отличие генетической рекомбинации от всех типов мутации, рассмотренных выше состоит в том, что в результате ее в наследственную ДНК бактерии извне привносится ранее отсутствовавший в ней блок генетической информации в виде совокупности новых для бактерии генов. Забегая вперед, следует сказать, что из трех способов возникновения рекомбинантов - трансформации, коньюгации и трансдукции наибольший интерес в рамках данной работы представляет трансдукция, так как этот процесс наглядно показывает, что ДНК вируса может встраиваться в ДНК бактерии без разрушения последней (по крайней мере в течение определенного времени).
Глава 3
Накопление полезных признаков
Базовая посылка эволюционных теорий видообразования в пользу постепенного формирования нового вида состоит в том, что в геноме особей исходного вида в результате мутаций появляются и под воздействием естественного отбора накапливаются полезные признаки.
Этот тезис вызывает у меня вопрос: каков критерий полезности вновь появившихся признаков?
Что касается человека, тут дело обстоит просто. Подходишь к такому носителю нового признака и задаешь вопрос в лоб: полезен ему этот вдруг появившийся у него признак или нет? И получаешь вполне определенный ответ. Но как быть со всеми остальными видами? Как ответить на этот вопрос не впадая в антропоцентризм? С точки зрения человека увеличение урожайности пшеницы после воздействия на ее семена гамма-излучения признак несомненно полезный, но вот с точки зрения пшеницы все может быть не столь однозначно.
Решение любого вопроса должно начинать с задания достаточно корректных начальных условий. Я предлагаю довести ситуацию до предельного состояния и посмотреть, как будет обстоять дело, если некий приобретенный в результате мутации признак в связи с изменением условий существования вида вдруг становится абсолютно необходимым для выживания вида. Другими словами в геноме особи есть пара комплементарных генов, представленных в виде доминантного (А) и рецессивного (а) аллеля. Интересующий нас признак контролируется одним из этих аллелей. В этих же начальных условиях оговариваем, что в качестве исходного материала у нас есть две особи противоположного пола - носители указанной пары генов. Жизненную задачу этих особей - произвести на свет носителей любой возможной комбинации этих генов тоже считаем реально выполнимой.
Тот факт, что мутация гена, приведшая к возникновению нового признака, может произойти только у одной особи нас смущать не должен. Допускаем, что особь эта после оной мутации находилась в репродуктивном возрасте и последующие поколения унаследовали бесценный дар.
Результаты усилий нашей пары производителей приведены в таблице.
|
|
Аа |
|
Аа |
АА Аа Аа аа |
Для простоты рассуждений было принято, что потомство взятых нами производителей включает по одной особи с данным сочетанием аллелей. Наличие двух особей с сочетанием аллелей -Аа- обясняется тем, что аллели получены от разных родителей.
При условии, что абсолютно необходимый для выживания признак контролируется доминантным аллелем (А), признаем особь с сочетанием аллелей -аа- нежизнеспособной и считаем, что она отбудет в мир иной до достижения репродуктивного возраста.
Отметим также тот факт, что в первом поколении появится особь с мономорфным (гомозиготным) сочетанием аллелей (АА) по рассматриваемому признаку и гомозиготность по этому признаку в целом в первом поколении среди достигших репродуктивного возраста особей составит 33%..
Таким же образом составляем таблицу для второго поколения, Как и в первом случае учитываем наличие мужской и женской особи, несущих одно и то же сочетание аллелей.
|
|
АА |
Аа |
Аа |
|
АА |
АА АА АА АА |
АА Аа АА Аа |
АА Аа АА Аа |
|
Аа |
АА АА Аа Аа |
АА Аа Аа аа |
АА Аа Аа аа |
|
Аа |
АА АА Аа Аа |
АА Аа Аа аа |
АА Аа Аа аа |
Всего получено 36 вариантов сочетаний из них 4 составляют сочетание -аа-. Но наряду с этим стоит обратить внимание на то, что процент гомозиготности аллелей по рассматриваемому признаку у жизнеспособных особей во втором поколении вырос и составил уже 50%.
Таблица для третьего поколения имеет размер 32 х 32 (32 строки и 32 столбца), что не позволяет включить ее в текст, да это и не имеет особого смысла. Проще определить количество всех сочетаний и определить процент мономорфности -АА- математическим путем.
Вот данные для третьего поколения.
Общая численность потомства 4096 особей.
Нежизнеспособных особей (несущих в геноме сочетание -аа-) 256.
Процент гомозиготности аллелей по рассматриваемому признаку - 56,25%.
Мы снова видим увеличение процента гомозиготности по рассматриваемому признаку и обоснованно можем предположить, что этот процент, хотя и значительно медленнее, будет расти и далее.
Рассмотрим теперь ситуацию, когда жизненно необходимый для выживания признак контролируется рецессивным аллелем. Здесь ситуация намного проще. Из первого поколения выживет лишь одна особь с сочетанием аллелей -аа-, т.е. процент гомозиготности по рассматриваемому признаку сразу станет равным 100% и таковым останется и в последующих поколениях. При условии, конечно, что изначально популяция располагает достаточным количеством носителей сочетания аллелей -Аа-. Но с этим особых проблем не просматривается, особенно если с момента возникновения мутации, вызвавшей появления аллеля -а- до момента когда его наличие в геноме особей стало жизненно необходимым сменилось достаточное количество поколений, что опять же не невероятно, учитывая, что в природе изменение условий обитания вида чаще всего происходит достаточно плавно.
Мутации, как известно, могут привести к появлению не только полезных, но и вредных признаков. Но нетрудно заметить, что вредный признак - это тот же полезный признак, но со знаком минус. Таким образом, ситуация с абсолютно вредными признаками сводится к уже рассмотренной.
Вернемся к тому, что мы рассматривали процесс накопления признаков в ситуации предельной, вероятность которой в природе пренебрежимо мала. Основная часть мутаций приводит к появлению относительно (в сравнении с имевшимся ранее) вредных и относительно полезных признаков. Но достаточно обоснованно можно предположить, что несмотря на это тенденции, выявленные выше, будут иметь место и в этом случае. Таким образом можно сделать предварительный вывод.
Полезные в той или иной мере для выживания вида (популяции) признаки, возникшие в результате мутаций, сохраняются в геноме этого вида (популяции). C течением времени происходит увеличение процента мономорфности (гомозиготности в рамках всей популяции) по этим признакам, достигающего при некоторых условиях 100%.
Как нетрудно заметить, до сих пор все результаты наших рассуждений полностью подкрепляли позиции эволюционных учений. Особо выделен лишь факт наличия мономорфности аллелей, контролирующих полезные для выживания вида признаки и перманентного увеличения процента этой мономорфности (гомозиготности по этим признакам в рамках популяции).
Но пойдем дальше.
Для того, чтобы процент мономорфности возникшего в результате мутации аллеля - носителя полезного признака - имел тенденцию к перманентному росту, полезность кодируемого им признака, как было сказано выше, должна быть тесно связана с вновь приобретенным (в результате той же мутации) преимуществом особи в выживании. А теперь зададимся вопросом: а может ли мутация одного гена сразу дать искомое преимущество? И вспомним при этом какое количество генов включает в себя даже простейший геном, а также то, что зачастую признак контролируется далеко не одним геном.
Кроме того любой живой организм является носителем не одного, а некоторой суммы признаков, следовательно, вполне возможно, что отклонение одного из них в положительную сторону может быть сведено на нет вариабельностью остальных. Точно так же возможен и противоположный эффект.
В результате, окончательный вывод о накоплении полезных признаков, возникающих в результате мутаций, должен быть примерно таким.
Полезность нового варианта любого признака, возникшего в результате мутации, всегда является относительной, не влияющей определяющим образом на выживаемость особи, вследствие чего она не выделяется естественным отбором и не приводит к увеличению процента мономорфности аллелей, контролирующих данный признак.
В случае же с вредными мутациями, дело обстоит несколько иначе. Если вредоносность мутации незначительна, она сохранится в геноме. Но если вредоносность мутации превзойдет некоторый порог, особь будет неминуемо отбракована естественным отбором.
Глава 4
Признаки видовые и индивидуальные
Представим себе самую обычную ситуацию: в семье родился ребенок.
Счастливый папа через несколько дней доставляет домой маму и дитя. Начинаются визиты бабушек и дедушек, дядей и тетей, сестер и братьев, знакомых и соседей. Главное, что их интересует - на кого же ребенок похож? Вот уши у него явно папины, а губы - мамины. Нос как у дедушки, а разрез глаз как у бабушки. А вот со цветом глаз что-то не получается. Но это никого не смущает - знать так наследственность распорядилась - может у прадедушки были глаза такого цвета.
Ситуация, как видим, самая обычная, и каждый из нас не раз был ее свидетелем. Но есть в ней одна деталь. Никому не кажется странным, что ребенок - это человек. У него две руки и две ноги, два глаза и два уха, на руках и на ногах по пяти пальцев - словом у него присутствует все, что делает его человеком, у него присутствуют все признаки, характеризующие человека как биологический вид.
То же самое относится ко всему живому на земле. Беглого взгляда достаточно нам, чтобы сказать: "Это собака. Это кошка. Это таракан. Это сосна. Это роза. Это карась."
При таком отнесении живых существ к определенному виду нам неважно, что эта конкретная сосенка немного кривовата, и что у одной собаки уши стоят торчком, а у другой повисли лопухами. Собака остается собакой, а сосна - сосной, и признаки, которые свойственны именно этой собаке и именно этой сосне в плане определения принадлежности к виду во внимание обычно не принимаются. Точно так же пусть далеко не все наши знакомые на одно лицо, а скорее наоборот, пусть в масштабах села, города, да что там мелочиться - в мировом масштабе трудно найти двух совершенно схожих между собой людей (если только они не однояйцовые близнецы), это не мешает им оставаться людьми.
Любой представитель животного или растительного вида на Земле относится к какому-то биологическому виду, и несет в себе все характерные признаки этого вида -видовые признаки. И глядя на что-то живое мы в первую очередь определяем это живое как вид.
Ну а поскольку наличие любого, присущего данной особи, признака обусловлено генетически, то вполне справедливым будет утверждение, что присутствие у любой особи, принадлежащей к данному виду, всех признаков, характеризующих этот самый вид, обусловлено наличием в геноме каждой особи общей для всех особей данного вида совокупности генов, контролирующих эти признаки.
Именно эта совокупность генов (назовем ее видоопределяющей) определяет, что у человека должно быть два глаза, две руки, две ноги, что большой палец на ладони у человека противопоставлен остальным четырем пальцам и т.д. Именно видоопределяющая совокупность генов определяет строение и функционирование общих для всех людей органов и систем - нервной, мышечной, кровеносной, лимфатической и других. Только благодаря наличию этой совокупности генов существует, например, медицина, основанная на изучении и лечении общих для всех людей патологий имеющихся у каждого человека органов и систем.
Наличие в геноме видоопределяющей совокупности генов - факт объективный, ибо в противном случае отвергается и другой объективный факт - факт существования биологического вида.
Гамма видообразующих признаков может быть определена на основе некоторой выборки особей.
Разумеется, размер выборки для определения совокупности видовых признаков данного биологического вида должен обеспечивать получение достоверного результата. Но не менее важным фактором при составлении такой выборки является наличие в ней особей, принадлежащих к разным популяциям этого вида. Если, например, определять видовые признаки человека на основании выборки составленной из народов Юго-Восточной Азии, то можно сделать заключение, что одним из таких признаков является наличие на голове прямых черных волос. На основании же выборки, составленной из народов Африки, можно прийти к выводу, что видовым признаком человека является наличие черных курчавых волос. И только при взаимном сравнении полученных данных можно сделать правильный вывод, что видовым признаком человека является само наличие волосяного покрова на голове (во всяком случае до определенного возраста).
Поскольку видовые признаки, наличие которых у любой биологической особи обусловлено видоопределяющей совокупностью генов, одинаковы у всех особей, принадлежащих к данному виду, предварительно можно предположить, что и гены видоопределяющей совокупности мономорфны у всех особей данного вида. Но сразу необходимо оговорить, что, как будет показано далее, мономорфность генов видоопределяющей их совокупности не связана с проблемой выживания в том плане, как это было рассмотрено выше, а обусловлена иными причинами.
Аналогично тому, как это было сделано для видоопределяющей совокупности генов, в геноме любой особи может быть выделена еще одна совокупность генов - совокупность, отвечающая за индивидуальные признаки особи. Легко усматриваемое многообразие признаков одного и того же органа, варьирующееся от одной особи к другой - отсутствие мономорфности контролирующих эти признаки аллелей - говорит о том, что связь индивидуальных признаков с вопросами выживания достаточно косвенна.
Нетрудно заметить, что индивидуальные признаки некоторым образом вторичны, так как представляют из себя, если так позволено будет выразиться, признаки признаков, т.е. определяют второстепенные характеристики признаков первичных - видовых.
Совокупность генов несущая информацию об индивидуальных признаках, определяет признаки характерные именно для данной особи. Эта совокупность, если опять говорить о человеке, содержит информацию о форме носа, ушей, цвете волос и глаз, группе крови и т.д.
Изменение в результате мутаций генов, составляющих вторую их совокупность в геноме, контролируемых ими признаков, может быть достаточно радикальным, видимым даже невооруженным глазом, но, как уже было отмечено, не сопровождаться мономорфностью контролирующих их аллелей. Одной из причин, по которой это происходит, является то, что (этот момент также был уже отмечен) далеко не все индивидуальные признаки присущие особям данного вида имеют отношение к проблеме выживания в данных конкретных условиях. Так, например, мутация гена, контролирующего цвет глаз не сказывается на способности видеть. То, что мочки ушей могут быть или приросшими или не приросшими, никак не сказывается на функционировании слухового аппарата.
Наличие же рас у людей, равно как и отличия в фенотипе у различных популяций особей одного вида в то же время показывает, что определенные индивидуальные признаки дают искомое преимущество при выживании в определенных условиях. Об этом еще будет говориться позднее.
Из-за полиморфности генов, составляющих рассматриваемую совокупность чаще всего невозможно сказать, глядя на цвет глаз у папы и мамы, какой цвет глаз будет у их сына. Это происходит потому, что данная совокупность генов содержит информацию не только о признаках, проявленных у данной особи, но и о скрытых признаках - о тех признаках, которые могут появиться у потомства этой особи.
Чем еще отличаются между собой гены, образующие видоопределяющую совокупность, и совокупность, определяющую индивидуальные признаки особи?
Они отличаются по степени взаимосвязанности контролируемых ими признаков с другими признаками.
Мутационное изменение индивидуального признака или ничего не меняет во взаимной связанности органа, к которому относится этот признак, с другими органами и системами, либо такие изменения пренебрежимо малы. Как может сказаться изменение формы носа на его иннервировании и кровоснабжении? Как может повлиять на железы внутренней секреции изменение разреза глаз или вариации в цвете кожи?
Изменение же даже на уровне одного гена в видовом признаке, например, в строении какой-либо мышцы (именно в строении, а не в размерах, скажем) очевидно требует согласованного изменения в нервной и кровеносной системе, обеспечивающих функционирование этой мышцы. Что уж говорить в том случае, когда в процессе эволюции (согласно эволюционным теориям) происходит формирование нового видового признака, как то: электрических органов у угрей и скатов или ядовитых желез у змей.
Вот это отличие между генами, контролирующими видовые и индивидуальные признаки, представляется мне не просто важным, но очень важным.
Но как быть с тем фактом, что в общем-то все без исключения гены, образующие геном биологической особи, в той или иной степени связаны с наличием, строением и функционированием присущей этой особи органов и систем? Можно ли в таком случае даже условно выделять в геноме его видообразующую часть и часть, ответственную за индивидуальные признаки? Вопросы эти вполне закономерны, но приведенная выше аргументация делает такое выделение, на мой взгляд, вполне обоснованным.
Границу между видовыми и индивидуальными признаками в некоторых случаях провести достаточно сложно, но эта часть вопроса и не является предметом рассмотрения в настоящей работе. Кроме того "Третий постулат" представляет из себя всего лишь каркас, который нуждается в наполнении детально разработанным содержанием. Принимая во внимание, что в рамках развития эволюционного подхода к видообразованию работала не одна тысяча специалистов, сравниваться с которыми по квалификации для меня было бы явным проявлением мании величия, я и не претендую на тщательную проработку всех возникающих вопросов, а просто придерживаюсь принципа, высказанного О. Бендером: "Главное - это плодотворная дебютная идея."
Написал эту фразу и подумал: назовут ведь теперь меня эволюционисты "Бендером от биологии", определенно назовут. С них станется...
Глава 5
Накопление полезных признаков в плане возникновения нового вида
Вероятность мутации, перестраивающей ген, таким образом, что он становится носителем признака более полезного, чем тот, который был закодирован в нем до этой перестройки сама по себе не просто мала, она ничтожно мала по той же причине, по которой не стоит надеяться на то, что случайным образом перемешанный осмысленный текст превратится если не в литературный шедевр, то хотя бы в творение графомана. Что же говорить о вероятности мутаций, приводящих к появлению нового видового признака?
Тезис о возникновении нового вида в результате накопления полезных признаков, возникших в результате генных мутаций в видоопределяющей части генома, мягко говоря не имеет под собой основы по причине полной невозможности этого процесса. В самом деле, гипотетически полезное закрепление результатов мутации генов, входящих в видоопределяющую часть генома и контролирующих какой-либо видовой признак, просто напросто невероятно из-за требуемой цепи согласованных изменений в связанных с изменившимся в результате мутации органом других органов и систем. Причем все изменения во взаимосвязанных органах и системах, возникающие в результате совершенно случайных ошибок при репликации генов, совершенно случайных спонтанных мутаций и совершенно случайных воздействий мутагенных факторов должны происходить в требуемом направлении и абсолютно синхронно.
Я задаю простейший вопрос. Какие примеры могут служить подтверждением факта формирования у особей какого-либо вида нового видового признака, что в конечном счете приведет к возникновению нового вида? У того же человека, несмотря на то, что численность людей на Земле измеряется уже миллиардами?
Похоже, что мой вопрос является риторическим.
Опять же как быть с близкородственным скрещиванием?
Поясняю, что я имею в виду.
Законодательством многих стран, в том числе и России, запрещены браки между людьми, связанными определенной степенью родства. И это не чья-то прихоть. Этот запрет вызван здравым осознанием того факта, что близкородственные браки значительно чаще чем "обычные" становятся причиной появления потомства с врожденными уродствами.
А ведь такие браки могли бы служить прекрасным подтверждением тезиса о накоплении полезных признаков. Ведь даже будь эти признаки спрятаны в рецессивных аллелях, то где как не в близкородственных браках они могли бы явить себя миру? Однако же нет ни одного примера проявления в результате близкородственного скрещивания чего-то очень полезного, все обстоит как раз наоборот. И не только у людей. Все породы собак произведены от ограниченного числа производителей другими словами в результате близкородственного скрещивания, и всем породам свойственна пониженная жизнеспособность - не зря ведь в космос первыми летали не ротвейлеры, скажем, а обычные подобранные на улице дворняжки. То же можно сказать и о гомозиготных линиях лабораторных мышей. В общем, за примерами далеко ходить не надо.
Результаты близкородственного скрещивания говорят и о том, что тезис, который я отстаивал в предыдущих редакциях "Третьего постулата" о стопроцентной мономорфности видоопределяющей совокупности генов (гомозиготности этой совокупности в границах популяции) к моему глубокому сожалению неверен.
Тем не менее, доказательная часть работы от признания этой моей ошибки не требует пересмотра, сколь ни странным может показаться это утверждение. Просто вопрос о наличии или отсутствии мономорфности генов в видоопределяющей части генома нужно рассмотреть внимательнее.
Как ни странно, справедливость доказательной части работы обосновывает то же близкородственное скрещивание, ведь результат его - появление на свет особи (пусть и дефектной) того же вида, к которому принадлежат ее родители. Дефектность потомства, полученного в результате близкородственного скрещивания, объясняется тем, что мутации в видоопределяющей совокупности генов, в результате которых искажение контролируемого мутировавшим геном признака перешло некоторую границу, нарушают взаимную связь видовых признаков, так как в этом случае изменение, допустим, строения какого-либо органа не сопровождается соответствующим изменениями в связанных с этим органом иных органах и системах. Отсюда следует, что мутационные изменения в видоопределяющей совокупности генов допустимы в очень узких пределах, не затрагивающих взаимосвязанность видовых признаков, и гены этой совокупности в норме остаются очень близки к своей "дикой" - домутационной - форме. Данное обстоятельство позволяет считать гены этой совокупности независимо от величины процента их мономорфности моноинформативными.
Таким образом, несмотря на наличие в видоопределяющей части генома рецессивных аллелей, эта часть генома моноинформативна, то есть информация, хранимая генами этой совокупности, может изменяться без угрозы для существования особи и вида в целом в очень небольших пределах.
Глава 6
Селекционная работа, изменение условий обитания, естественный отбор и возникновение нового вида
Усилия селекционеров, на протяжении тысячелетий занимающихся выведением новых собачьих пород дали поразительные результаты - стоит посмотреть на стоящих рядом сенбернара и тойтерьера. Но один результат так и не был достигнут. Новый вид не возник. Кроме того, тот же опыт селекции на этот раз в деле возвращения к жизни вымерших зубров показывает, что, грубо говоря, используя генетический материал сенбернаров ли, тойтерьеров ли (в крайнем случае их обоих вместе с добавкой еще какой-нибудь породы) вполне возможно возродить исходного прародителя собак вообще.
Необходимо, однако, заметить, что процесс возрождения зубров не есть процесс восстановления исчезнувшего вида, т.к. этот процесс шел путем восстановления фенотипа зубра путем полового скрещивания особей, сохранивших признаки этого фенотипа. На каждой ступени этого восстановления полученное в результате потомство сохраняло возможность также произвести в свою очередь потомство и потомство плодовитое, следовательно есть достаточные основания полагать, что и восстановленные зубры способны к скрещиванию и производству плодовитого потомства с особями того вида, что был использован для процесса восстановления. А это как раз и свидетельствует о правильности сделанного замечания. В свою очередь этот факт свидетельствует о том, что или зубры изначально не представляли из себя самостоятельного вида, как не представляют из себя самостоятельных видов сенбернар и тойтерьер, или восстановлена лишь внешняя копия вида действительно существовавшего и впоследствии исчезнувшего.
В чем, на мой взгляд, причина невозможности создать новый вид путем селекции?
Дело в том, что селекционер в своей работе может путем отбора производителей воздействовать только на совокупность генов, контролирующую индивидуальные признаки особи.
Именно за счет изменения состава этой совокупности генов, возможность которого обеспечивает наличие доминантных и рецессивных аллелей, и происходит выведение новых пород животных и сортов растений.
Видоопределяющая же совокупность генов моноинформативна. А поскольку усилия селекционера не могут ни изменить хранящуюся в генах информацию, ни, тем более, внести информацию новую, то видоопределяющая часть генома абсолютно устойчива к усилиям селекционера и селекционная работа к появлению нового вида не может привести в принципе.
Если селекционная работа чем-то отличается по своему воздействию на генетическую информацию, хранимую каждой особью вида, от воздействия изменения условий обитания, пойдем в природу. Но и в этом случае придется признать, что ни изменение условий существования ни естественный отбор точно так же как и селекционный процесс не способны ни изменить хранящуюся в генах информацию ни внести в исходный геном все ту же новую генетическую информацию.
Если даже для рассматриваемого случая допустить ранее отвергнутую возможность накопления полезных признаков в видоопределяющей части генома, при отсутствии целенаправленных усилий селекционера, работающего над каждым поколением тех же собак, процесс проявления у собачьего рода новых признаков и гипотетически возможного возникновения нового вида в природных условиях растянется на невообразимо длительный срок. За это время, вдобавок ко всему, не один раз сменятся условия обитания вида, и даже один этот фактор, не мешая при каждом таком изменении проявлению и закреплению в потомстве полезных для выживания признаков, делает вообще сомнительным образование нового вида эволюционным путем. И в самом деле: стоит только виду явить ряд признаков несомненно полезных в новых условиях, но не приведших еще к образованию нового вида, как условия существования вполне могут снова измениться, вследствие чего может оказаться, что вновь приобретенные признаки являются в лучшем случае бесполезными, все начнется сначала, и, таким образом, процесс образования нового вида под воздействием изменения условий существования грозит стать бесконечным.
Положения вещей не меняет явление кроссинговера, заключающееся в обмене между хромосомами участками, несущими аналогичную информацию. Неравный кроссинговер, позволяющий увеличить количество генов в геноме особи, и, следовательно, формально способный повлиять на имеющуюся в геноме информацию о видовой принадлежности особи при том условии, что он происходит в видоопределяющей совокупности генов принципиально не может ничего изменить в факте принадлежности данной особи к конкретному виду вследствие моноинформативности этой совокупности. Неравный кроссинговер в этом блоке наследственной информации в лучшем случае может лишь исказить затронутый им видовой признак или элементарную его составляющую, но никак не вызвать появление нового видового признака. (Напрашивающаяся в данном случае аналогия: случающееся повторение страниц в книжном тексте ничего не добавляет к его содержанию.)
Эти же соображения относятся и к любым другим процессам, происходящим внутри генома, поскольку эти процессы также происходят в рамках уже имеющейся в геноме информации и в лучшем случае могут только произвести рекомбинацию этой информации.
Геном особи в данной работе рассматривается, как нетрудно заметить, в первую очередь, как информационная система. И в этом плане любой ген для меня, как для инженера, это в первую очередь не часть биологической структуры, оформленной в виде хромосом, а пакет информации, которой надлежит быть реализованной в процессе развития и жизнедеятельности живого организма.
И еще раз следует сказать, что не всякое воздействие на носитель информации является информационным воздействием. На мой взгляд понять это может помочь простая аналогия. Если в бензобаке вашего автомобиля кончается бензин, то для того, чтобы восполнить его запас, вы едете на ближайшую бензоколонку, а не принимаетесь стучать молотком по крышке бензобака. Равным образом рекомбинация имеющейся информации не есть внесение новой информации, Вероятнее всего любая рекомбинация - это внесение искажений в информацию, имеющую определенный смысл. Аналогия более широкого плана: удар молотком (неинформационное воздействие) по сложенному в определенном порядке набору детских кубиков, на каждой стороне которых изображен фрагмент картинки (набор кубиков здесь - пакет информации), скорее всего не заставит кубики сложиться так, чтобы образовалась новая картинка. Если же добавить к имеющимся кубикам новые кубики с новыми фрагментами рисунка (информационное воздействие), то шансы получить более содержательное изображение существенно возрастут.
Здесь уместно сделать некоторое отступление. Закономерен вопрос: как же тогда в принципе появились рецессивные аллели? Ведь происхождение их в результате мутаций является наиболее вероятным. А это значит, что в результате мутаций возникновение новой генетической информации возможно. На мой взгляд факт наличия в геноме рецессивных аллелей объясняется следующим образом. Если представить живой организм как систему, обрабатывающую в процессе эмбриогенеза и онтогенеза некоторую генетическую информацию, то он способен обработать эту информацию в некоторых пределах, то есть "полоса" обрабатываемой организмом генетической информации имеет некотрую "ширину". При этом условии живой огранизм способен обработать и то искажение исходной генетической информации, которое несет рецессивный аллель, придавая, таким образом, этому искажению статус новой генетической информации.
Еще раз следует сказать, что вышесказанное относится только к аллелям, контролирующим индивидуальные признаки, что следует из уже сказанного о видовых и индивидуальных признаках. Но это не умаляет значения того факта, что возникновение новых аллелей, контролируюших индивидуальные признаки, расширяет возможность приспособляемости живых организмов к меняющимся условиям обитания и способствует выживанию биологического вида.
Если же все-таки согласиться по методу доказательства от обратного с тем, что изменение условий обитания в совокупности с естественным отбором или работа селекционера способны вызвать появление нового биологического вида, то необходимо признать, что в геноме вида-предшественника уже содержатся все признаки нового вида - и видовые и индивидуальные. В самом деле - если их там нет, то как может привести изменение условий обитания и селекция, не воздействующие на структуру генов и не вносящие в геном новой генетической информации, к их появлению, и откуда тогда взяться новому виду? Ведь из цилиндра можно вынуть кролика только если перед этим его туда посадить, как бы ни клялись зрители, наблюдающие за этим фокусом, что никто кролика в цилиндр не сажал, а появился он там сам собой. Рассматривая же в свете сделанного выше допущения процесс эволюции в ретроспективе, не остается ничего другого, как признать, что ДНК первой на Земле живой клетки уже содержала в себе всю информацию, необходимую для появления "венца творения".
Сказанного достаточно, чтобы сформулировать первый постулат предлагаемой новой теории видообразования.
Первый постулат: возникновение нового вида возможно только при изменении содержащейся в геноме вида-предшественника информации о видовой принадлежности особей, его составляющих.
Глава 7
Объем видоопределяющей части генома и возникновение нового вида
Как уже было сказано, внешние мутагенные факторы, если мы признаем за ними способность к целенаправленному изменению генетической информации действуют в рамках неизменного объема генетического материала, так как не привносят в геном "строительный материал" для "постройки" новых генов. А из этого следует, что в результате воздействия мутагенных факторов обязательно должно происходить стирание или существенное искажение информации ранее закодированной в мутировавших генах.
При этом становится совершенно непонятным, почему тот же человеческий эмбрион при своем развитии являет в своем строении признаки биологических видов, предшествовавших ему на ступенях эволюционной лестницы, ведь, как только что было сказано, сведения об этих ступенях в ДНК эмбриона должны были или исчезнуть вообще, или оказаться существенно искаженными. А между тем наличие в геноме информации о видовых признаках видов-предшественников принципиально необходимо для развития эмбриона. Как невозможно, снова переходя к аналогиям, построить первый этаж здания, не выстроив предварительно фундамент, а второй этаж - не построив первый, так и эмбрион в процессе своего развития до появления на свет должен повторить эволюционный путь вида, так как каждая предыдущая ступень в процессе развития эмбриона соответствует определенной ступени на пути эволюции вида и является фундаментом для последующей.
На каждой предыдущей ступени происходит накопление биологического материала (клеток, зачатков органов, тканей), являющегося основой для частичного или полного использования генетической информации, соответствующей следующей ступени развития эмбриона. В противном случае эмбрион на любой стадии развития представлял бы из себя всего лишь уменьшенную копию полностью сформировавшейся особи, что свидетельствовало бы только о том, что ни о какой эволюционной преемственности при возникновении нового вида говорить не приходится.
В дополнение можно сказать, что если рассмотреть процесс возникновения нового вида в результате генных мутаций в свете эволюционных учений в ретроспективе, то окажется, что он базируется на том условии, что объем генома одинаков у, допустим, человека, и у амебы (если она была эволюционным предшественником человека), что при всем моем уважении к амебе я не могу не считать абсурдом.
Процесс возникновения нового биологического вида под воздействием мутагенных факторов имеет и другие следствия дополнительно обосновывающие его невозможность, но эти следствия будут рассмотрены ниже. Сказанного же достаточно, чтобы сформулировать второй постулат.
Второй постулат: возникновение нового биологического вида возможно только при условии увеличения объема информации, содержащейся в видоопределяющей части генома вида-предшественника и, следовательно, при условии увеличения размеров этой части генома.
Второму постулату можно поставить в соответствие тот факт, что описание более сложного объекта содержит больше информации, чем описание объекта более простого. Но из этого следует, что увеличение размеров видоопределяющей части генома или, что то же самое, количества информации о видовой принадлежности особи ведет к возникновению вида более продвинувшегося по пути эволюции.
Верно и обратное утверждение: для продвижения вида по пути эволюции необходимо увеличение количества информации, содержащейся в блоке информации о видовой принадлежности и, следовательно, размеров этого блока.
Два эти следствия связаны теснейшим образом и, взятые в совокупности, объясняют, почему видообразование движется по пути от простого (клетка) к сложному (допустим - человек). Действительно, возьмем в качестве точки отсчета вид, стоящий на определенной ступени эволюционной лестницы и соответствующий этой ступени объем видоопределяющей совокупности генов, характеризующей этот вид. После этого начнем описание процесса эволюции этого вида с нуля, заполняя этот объем новым содержимым и используя при этом тот же генетический код. В конечном итоге мы, возможно, получим "описание" нового вида, но по всей совокупности признаков - по сложности своей - он окажется на той же ступени эволюции, что и вид, взятый в качестве точки отсчета. Таким образом, сохранение объема видоопределяющей совокупности генов неизменным означает прекращение процесса эволюции. При уменьшении же этого объема, в него возможно будет поместить только описание вида, стоящего на лестнице эволюции ниже исходного, что соответствует обращению процесса эволюции вспять.
Два рассмотренных варианта заполнения видоопределяющей совокупности генов новой информацией о видовой принадлежности возможно рассматривать только чисто теоретически. Невозможно начать процесс эволюции с чистого листа. Можно с помощью мутагенных факторов уничтожить часть генетической информации, сократив тем самым объем информации о видовой принадлежности, но это будет равносильно уничтожению вида. (Позднее будет рассмотрен теоретически возможный вариант сокращения информационного содержимого видоопределяющей совокупности генов, но с указанными уже последствиями). Остается единственный вариант - продвижение вида по лестнице эволюции выше и выше в результате увеличения количества информации о видовой принадлежности и, следовательно, увеличения объема видоопределяющей совокупности генов.
Глава 8
Предположение о существовании признакообразующих вирусов
Как уже говорилось, новые гены могут появиться в геноме, например, за счет неравного кроссинговера.
Но, вводя в рассмотрение этот факт, необходимо опять же допустить жесткое согласование появления нового гена за счет внутренних процессов в геноме и "перезаписи" его содержимого за счет воздействия факторов внешних, причем "перезаписи" именно в нужном направлении. Рассчитывать вероятность осуществления такого процесса представляется мне делом столь же полезным, как и споры древних теологов о том, сколько ангелов могут поместиться на кончике иглы.
Необходимость для возникновения нового вида внесения в геном вида-предшественника принципиально новой генетической информации напрочь отвергает возможность возникновения нового вида на пути манипуляций с генной информацией, уже имеющейся у особей, образующих вид-предшественник, равно как и с суммарным геномом популяции или вида-предшественника в целом. В подтверждение этого соображения можно привести примеры существования растений и животных (например, рыб) с полиплоидным набором хромосом. Подобные примеры могут служить лишь иллюстрацией многообразия процессов, происходящих в геноме, но никак не могут приблизить к пониманию того, каким образом возникает новый вид.
В поисках фактора, действие которого могло бы привнести в геном особи новую генетическую информацию, вспомним о вирусах. Факт, что вирусы могут содержать в себе ДНК, сходную по строению с ДНК, находящейся в хромосомах клеток зараженной вирусом особи, известен. Известно и то, что ДНК вируса может встраиваться в ДНК клетки, каковой факт был уже специально подчеркнут выше. Но, вместе с тем, известны и результаты вирусного поражения живых клеток.
Предположим, тем не менее, что существуют вирусы, которые могут встраиваться в ДНК половых клеток без их разрушения. При этом такие вирусы способны нести в своей ДНК ген, встраивание которого в ДНК клетки означает, что отныне ДНК этой половой клетки содержит в себе информацию о новом элементарном признаке, контролируемом этим геном.
Механизм действия таких вирусов, вероятнее всего, должен быть следующим. Они (вирусы) должны или поражать репродуктивные органы, заставляя их продуцировать половые клетки, несущие информацию о новом элементарном признаке, или же, резидентно присутствуя в клетках репродуктивных органов, они должны перманентно поражать созревшие половые клетки без их разрушения. В любом из случаев обретенный половыми клетками в виде нового гена новый элементарный признак проявится в потомстве инфицированных особей.
Однако, рассмотрим, к каким следствиям ведет допущение существования признакообразующих вирусов.
Вполне очевидно, что любой видовой признак является сложным, состоящим из множества элементарных, следовательно, его наличие контролируется не одним геном, но только их совокупностью.
Понятно также, что признакообразующие вирусы не могут действовать согласованно, следовательно, процесс возникновения нового вида будет под их воздействием происходить ступенчато, причем каждая пройденная ступень соответствует приобретению видом нового элементарного признака.
Пусть однократное воздействие вируса привело к появлению у потомства инфицированной особи появление элемента нового видового признака.
Заражение признакообразующим вирусом происходит в рамках всей популяции, что представляется более вероятным, чем любое другое течение событий, то есть такое заражение имеет характер эпидемии. В этом случае нет трудностей с распространением нового элементарного видового признака, так как составляющие популяцию особи, сохраняя в себе признакообразующий вирус в резидентной форме способны производить только потомство, обладающее новым элементарным видовым признаком.
Но для образования полноценного нового видового признака необходимо последовательное и упорядоченное воздействие на популяцию череды признакообразующих вирусов, каждый из которых привносит элементарную составляющую этого признака, ибо только по завершении этого процесса популяция станет новым видом.
Поэтапное формирование нового вида уже рассматривалось в привязке к мутациям. Так что основные черты этого процесса, приводящие к его принципиальной невозможности уже известны.
Но допущение существования признакообразующих вирусов снимает одну из проблем - проблему внедрения в геном новой генетической информации, то есть внедрение в геном признакообразующих вирусов - это информационное воздействие на геном. В связи с этим имеет смысл вновь пройтись тернистым путем поэтапного формирования вида, но с признакообразующим вирусом в качестве поводыря.
Предположение о возможности возникновения нового вида путем последовательного накопления элементарных видовых признаков под воздействием признакообразующих вирусов, влечет за собой по крайней мере три следствия (которые в равной мере относятся и к процессу возникновения нового вида под воздействием мутагенных факторов).
Во-первых, понятие "биологический вид" при таком способе возникновения нового вида теряет всякий смысл, ведь отличие в один элементарный видовой признак между двумя сменяющими друг друга поколениями особей этого вида это даже не отличие между особями, принадлежащими к близкородственным видам, - это отличие неимоверно меньших размеров. Большего же различия между двумя следующими друг за другом, условно говоря, "промежуточными видами" добиться на таком пути возникновения нового вида невозможно. Сам же процесс видообразования при этом становится непрерывным и бесконечным, ведь стоит закончиться формированию одного нового видового признака, как тут же может начаться формирование признака следующего и т.д., и т.д.
Во-вторых, невозможно представить, как будет выглядеть приобретение видом нового видового признака путем накопления элементарных составляющих этого признака.
В-третьих, при поэтапном формировании нового вида не должно быть никаких проблем с обнаружением ископаемых останков "переходных звеньев", собственно, ими должны в таком случае стать все ископаемые останки, что, как известно, в действительности места не имеет.
Но вследствие предположения о существовании признакообразующих вирусов в конце тоннеля появился свет!
Наступило время свести воедино все сказанное выше. Надеюсь достаточно
убедительно были доказаны следующие положения:
- новый вид не может возникнуть в результате накопления признаков, возникших в
результате мутаций в видоопределяющей части генома;
- новый вид не может возникнуть в результате воздействия на блок наследственной
информации об индивидуальных особенностях особей вида;
- для возникновения нового вида генетическая информация о новом видовом признаке (новых
видовых признаках) должна быть занесена в блок наследственной информации, хранимый
видоопределяющей совокупностью генов вида-предшественника;
- занесение информации о новых видовых признаках в блок наследственной информации о видовой
принадлежности должно сопровождаться увеличением количества содержащегося в этом блоке
информации и связанным с этим увеличением размеров этого блока, что на практике означает
рост объема видоопределяющей совокупности генов, характеризующей вид-предшественник;
- увеличение объема видоопределяющей совокупности генов за счет мутаций не сопровождается
внесением в геном новой генетической информации и к возникновению нового вида привести не
может;
- возникновение нового вида вследствие увеличения количества информации в блоке
наследственной информации о видовой принадлежности, осуществляемого путем последовательного
занесения в него извне информации о каждом новом элементарном видовом признаке по
отдельности, невозможно.
Из всего вышевыделенного логично вытекает формулировка третьего постулата новой теории.
Третий постулат: изменение информации о видовых и индивидуальных признаках в геноме вида-предшественника для возникновения нового видового признака (признаков) и, следовательно, для возникновения нового вида должно происходить внесением в геном единого блока информации, при взаимодействии которого с наследственной информацией, присущей виду-предшественнику, возникают все видовые и индивидуальные признаки нового вида.
Глава 9
Основы новой теории
Предлагаемая теория видообразования основана на трех постулатах, обоснование и формулировка которых приведены выше, и двух теоремах, которым еще только предстоит быть доказанными единственно приемлемым в данном случае методом - практическим.
Теорема первая. Существуют некие организмы, назовем их видообразующими вирусами, способные нести в себе ДНК, содержащую объем информации достаточный для того, чтобы при соединении с наследственной ДНК пораженной вирусом особи модифицированная ДНК с момента такого соединения несла в себе все видовые и индивидуальные признаки нового биологического вида.
Теорема вторая. Видообразующие вирусы поражают в первую очередь, а может быть и исключительно, органы репродукции, причем таким образом, что мужские и женские половые клетки начинают нести в себе после инфицирования наследственную информацию, присущую уже новому виду.
Фундаментально предлагаемая теория возникновения нового биологического вида на основе третьего постулата базируется на неявном, но прямом следствии из второго начала термодинамики. Согласно этому следствию любая система, характеризуемая некоторыми параметрами, будучи изолированной по какому-либо из параметров от внешнего воздействия, может по этому параметру только деградировать.
Особо необходимо подчеркнуть, что будучи похожа внешне на существующие мутационные теории видообразования, предлагаемая теория коренным образом отличается от них внутренним содержанием.
Отличие состоит в том, что в предлагаемой теории новый вид возникает в результате внешнего информационного воздействия на геном вида-предшественника.
Нетрудно заметить, что при условии реальности существования видообразующих вирусов проблема происхождения новых видов в свете предлагаемой теории сводится к проблеме происхождения вирусов. Не только видообразующих, но вирусов вообще. Но это, как сказано у классиков, уже совсем другая история.
Глава 10
Размышления на тему
Теперь, когда сформулированы основные положения новой теории, прежде чем рассматривать процесс видообразования в ее свете стоит попробовать свести воедино все спорные места в эволюционных теориях видообразования, которые в конечном итоге и привели к появлению настоящей работы.
Для начала я попытаюсь еще раз представить процесс возникновения нового вида в свете эволюционных теорий.
Произведем мысленный эксперимент. Представим себе популяцию мышей, допустим, полевок, живущих на поле у опушки леса. Для чистоты эксперимента условимся, что и лес и поле принадлежат надежно изолированному для случайного проникновения на него любой другой живности, кроме уже имеющейся на нем, острову. Очень плавно, но неуклонно происходит изменение климатических условий, в результате которых ухудшается кормовая база полевок на том поле, на котором они живут и также медленно но неуклонно на деревьях, вблизи которых наши полевки проживают, растет урожайность очень подходящих для их питания семян и плодов. Эти семена и плоды никто не поедает, так что если мыши освоят эту новую кормовую базу, конкурентов у них не будет. Одна тонкость - эти семена и плоды съедобны только пока они не упали с деревьев. На острове имеется источник радиоактивного излучения, достаточно мягкого, чтобы только вызывать мутации в геноме наших мышей, имеет место факт наличия природных химических мутагенов, над островом регулярно возникают озоновые дыры, да и в норме солнечное излучение способствует этим самым мутациям. Космическое излучение, достигающее поверхности острова тоже имеет самые благоприятные кондиции. Внутренние процессы в геноме мышей протекают необходимым образом и тем самым вносят свою лепту в возникновение новых признаков. Условия заданы, и, на мой взгляд, являются идеальными для эволюционного возникновения нового вида мышей, способных беспечно резвиться на деревьях. Если называть вещи своими именами, то речь идет о превращении мышей в белок.
Я сильно подозреваю, что в конечном итоге для эволюционного подхода к проблеме и этого рая будет недостаточно, но совесть моя чиста - я сделал для наших мышей все, что мог.
Почему мыши изначально неспособны освоить новую экологическую нишу? Логично предположить, что это происходит потому, что присущий им опорно-двигательный аппарат в его исходном состоянии не позволяет им уверенно карабкаться и прыгать по стволам и ветвям и удерживаться на них при угрозе падения, кроме того строение зубов и челюстей также, мягко говоря, не способствует переходу на новую кормовую базу. Ну а если добавить, что определенные изменения должны произойти в строении органов пищеварения, центральной и периферической нервных систем, а также в железах внутренней секреции (в первую очередь тех, которые участвуют в пищеварении), то картина представляется довольно ясной.
Итак, считаем, что процесс пошел. Но остановим его на середине пути и вглядимся в его результаты. Что же мы увидим? Даже если изменения во всех органах и системах шли согласованно, то есть мутации происходили в нужное время и в нужных генах, что лично я представить себе не могу, то на момент оговоренной остановки мы имеем особь уже не способную жить в прежних условиях, но еще не способную существовать в условиях новых. Горька ее доля...
Кроме того, что значит тот факт, что мутации происходили согласованно? Это значит только то, что совершенно случайные по своей природе процессы приобрели упорядоченность и целенаправленность. Что придало им эти свойства? Трудно ожидать вразумительного ответа на этот вопрос.
Я могу представить с очень большими натяжками, что на пути постепенного формирования гаммы новых признаков возможно усложнение и совершенствование имеющихся у биологических особей органов и систем, но совершенно не могу представить возникновение на этом пути органов и систем новых, ранее отсутствовавших. Но, допуская возможность линейной эволюции, невозможно объяснить, как на таком пути из первой на Земле протоклетки возникло многообразие всего живого, что невозможно без разветвлений эволюционного ствола на отдельные ветви.
Но в общем-то вопрос накопления полезных признаков должен рассматриваться в еще более широком плане, чем это сделано выше.
Я задаю себе вопрос. Как может быть, что накопление признаков полезных для существования и выживания данного конкретного вида в данных конкретных условиях вдруг приводит к возникновению нового вида, способного существовать в условиях, не имеющих с исходными ничего общего? И не нахожу другого ответа кроме того, что тезис о накоплении полезных для выживания исходного вида признаков как непременного условия возникновения нового вида является ничем иным, как прямой подменой действительного желаемым.
Необходимо помимо всего прочего отметить, что до сих пор мною не было рассмотрено еще одно соображение. Возникновение нового вида обязано сопровождаться изменением кариотипа вида-предшественика. Кариотип (от карио... и греч tipos - образец, форма), совокупность признаков хромосомного набора (число, размер, форма хромосом), характерных для того или иного вида. Постоянство кариотипа каждого вида поддерживается закономерностями митоза и мейоза.
И далее. Изменение кариотипа может происходить вследствие хромосомных и геномных мутаций.(Определение взято из биологического энциклопедического словаря).
Это значит, что в тот момент, когда согласно эволюционным теориям количество накопленных полезных для возникновения нового вида признаков достигнет необходимой для возникновения нового вида величины, то есть по окончании принципиально невозможного процесса, должна вдобавок, как апофеоз, произойти еще и хромосомная или геномная мутация для возникновения кариотипа, характерного для нового вида.
Мутация, вызывающая изменение кариотипа, должна именно завершать процесс формирования нового видового признака (признаков). В противном случае надо допустить существование видов, обладающих абсолютно идентичным набором генов, но с разным кариотипом, что, в принципе, допустимо, если исходить из определения вида, которое будет приведено ниже, а также существование видов с недоформировавшимся видовым признаком (признаками), если стоять на позициях постепенного формирования этих новых признаков, а уж это допущение, по моему убеждению, выходит за всякие рамки.
Не знаю как у кого, но это немыслимое количество оговорок, натяжек и невыполнимых требований без которых возникновение нового вида эволюционным путем невозможно, вызывает у меня чувство умственной неполноценности, из-за неспособности создать логичную мысленную модель процесса эволюционного видообразования.
Чего стоит одна только необходимость для эволюционного видообразования последовательных и целенаправленных мутаций, вызываемых случайными факторами, причем мутаций, происходящих на протяжении жизни многих и многих поколений вида-предшественника. Позволительно спросить: кто и где хранит информацию о конечной цели мутаций, кто обеспечивает их согласованность и синхронность, кто определяет очередность мутаций и следит за ее соблюдением? Можно услышать в ответ, что конечно же никто этим не занимается, все происходит случайно, а вот естественный отбор и производит закрепление результатов мутаций, приводящих к возникновению нового вида.
Что можно сказать на это? Да и нужно ли?
При всем при этом эволюционисты в массе придерживаются атеистических взглядов, и всех имеющих неосторожность усомниться в истинности епэнэмэ (неологизм принадлежит В. Войновичу и означает ЕПНМ - единственно правильное научное мировоззрение) причисляют к креационистам, что конечно же делает абсолютно ненужными все дальнейшие разговоры. Какой с креациониста спрос?
Я не исповедаю какую-либо религию, на настоящий момент не принадлежу к какой-либо церкви. Но однажды и навсегда я решил для себя вопрос о существовании Создателя в положительном смысле. Тем не менее я считаю, что предлагаемый мной взгляд на проблему видообразования гораздо более материалистичен, чем взгляды эволюционистов. Во всяком случае до настоящего момента и, как это будет видно и в последующем, необходимость участия Творца в процессе формирования нового вида на основе третьего постулата не просматривается. Процесс достаточно убедительно описывается законами вероятности, что говорит о роли случайности в его осуществлении.
Но не случайно направление вектора эволюции - от первой клетки до человека разумного.
Глава 11
Видообразование по новой теории. Следствия
После заражения видообразующим вирусом популяции вида-предшественника потомство зараженной популяции являет собой новый биологический вид. Это значит, что процесс возникновения нового вида требует очень немного времени (он равен длительности репродуктивного периода одного поколения вида-предшественника), собственно, его можно назвать одномоментным. Популяция при этом выступает в роли видообразующей единицы. Кстати, именно одномоментность возникновения нового вида может служить, по крайней мере в некоторых случаях, объяснением обнаружения новых биологических видов на исследованных ранее территориях.
Лишенная возможности воспроизводства в первозданном виде популяция вида-предшественника вымирает. Но популяции того же вида достаточно удаленные от зараженной имеют реальную вероятность избежать заражения.
Таким образом, популяции вида-предшественника и вновь возникшего вида могут сосуществовать в течение длительного промежутка времени и ископаемые останки нового вида могут оказаться старше таких же останков вида, предшествовавшего ему на лестнице эволюции.
Сохранившиеся популяции вида-предшественника уже после появления на базе одной или нескольких из них нового вида продолжают оставаться базой для эволюции, так как сохраняется возможность заражения их новыми видообразующими вирусами. Таким образом, на базе одного вида-предшественника могут возникнуть разные виды.
На примере существования разных штаммов вируса гриппа можно сделать предположение, что исходные формы видообразующих вирусов также могут образовывать различные штаммы. В таком случае, если одна из популяций вида-предшественника была заражена одним штаммом видообразующего вируса, а другая популяция - другим, могут возникнуть близкородственные виды, например, осел и лошадь.
В том случае, если видообразующий вирус вирулентен для разных биологических видов, возможно возникновение двух или более новых видов, имеющих сходные видовые признаки (признак), например, сумчатые в Австралии.
Из новой теории видообразования следует, что наиболее совершенным в биологическом смысле вид является сразу после его возникновения. В самом деле, вновь возникший вид в рамках ограниченной по численности популяции сохраняет весь заложенный в его генофонде видовой и индивидуальный потенциал. Возможность проявления индивидуальных особенностей в неискаженной форме для каждой особи нового вида максимальна. Мутагенные факторы еще не успели оказать своего разрушающего воздействия и количество искажений наследственной информации, хранимой в ДНК особей нового вида, минимально.
Не может не обратить на себя внимание вывод о том, что процесс возникновения нового вида может считаться практически одномоментным и что новый вид появляется на свет биологически совершенным. Прямые аналогии с известным источником я комментировать не берусь.
После своего возникновения новый вид начинает осваивать подходящие экологические ниши, увеличивая ареал своего обитания и реализуя заложенный в его генофонде потенциал. Это время расцвета вида. Но одновременно с этим сразу после возникновения нового вида особи, принадлежащие к нему, начинают подвергаться воздействию всей гаммы мутагенных факторов, которые в полном соответствии со вторым началом термодинамики, если его применить к замкнутому объему генетической информации хранящейся в суммарном геноме нового вида, способны только разрушить или исказить наследственную информацию. Таким образом, расцвет биологического вида в том виде, каком он был обозначен выше, занимает достаточно ограниченное время, в то время как накопление искажений наследственной информации вследствие мутаций - процесс перманентный, и биологический вид с самого момента своего возникновения обречен на исчезновение.
Это утверждение становится тем более справедливым, если вспомнить, что новый вид возникает на базе наследственной информации вида-предшественника, уже несущей на себе груз накопленных искажений.
Что же происходит, если возникший вид вследствие его отличия от исходного вида окажется перед фактом отсутствия свободной экологической ниши? Он просто напросто исчезнет, не оставив никаких следов своего существования. Какова вероятность обнаружения останков считанного числа представителей нового вида, умерших в самом раннем возрасте?
Вновь возникший вид может стать исходным материалом для возникновения последующего вида, тем более, что возникновение этого последующего вида не означает исчезновение вида-предшественника, но не более того, и никак не может существовать, в течение неограниченного времени.
Из принципов, положенных в основу новой теории видообразования, прямо следует, что никаких "переходных звеньев" между видом-предшественником и вновь возникшим видом быть не может. Это, кстати, объясняет безрезультатность поисков "переходного звена" между обезьяной и человеком.
Тот факт, что в соответствии с новой теорией видообразования изменение наследственной информация о видовой принадлежности происходит путем ее "дописывания" означает, что существует принципиальная возможность, используя генетический материал некоторой выборки особей данного вида, восстановить методом клонирования весь эволюционный путь этого вида.
Этот же факт "дописывания" генетической информации объясняет сохранение в геноме вновь возникшего вида информации о видах-предшественниках, проявляемой в процессе развития эмбриона, что (и это было показано выше) не находит объяснения в свете эволюционных теорий.
Глава 12
Балластная часть ДНК. Последовательность реализации генетической информации в процессе развития эмбриона
Воздействие видообразующего вируса на клетку не уничтожает генетическую информацию, хранящуюся в ее ДНК, а лишь привносит новую. Логично предположить, что далеко не вся суммарная наследственная информация используется на пути превращения эмбриона в жизнеспособную особь, принадлежащую новому виду. Что стоит за этим предположением?
Как уже было сказано, наиболее полно должна использоваться информация о видовых признаках видов-предшественников. Таким образом, видоопределяющая совокупность генов должна включать в себя и гены, несущие информацию о видовых признаках видов-предшественников, без которой невозможно воплощение видовых признаков данного вида. Это объясняет наличие значительного количества одинаковых генов у представителей разных видов.
Тем не менее, нельзя исключить возможности того, что часть генетической информации о видовых признаках видов-предшественников в процессе эволюции становится ненужной. Тем более верным должно быть это предположение по отношению к индивидуальным признакам. В самом деле, информация о количестве, форме и расцветке чешуй кистеперой рыбы, которая может входить в ряд видов-предшественников человека, человеческой особи как представительнице вида не нужна. Мало того - в процессе эволюции эта информация, как и многая другая, лишилась базы для своего проявления. Это, в свою очередь, значит, что искажения ненужной генетической информации выпадают из-под санирующего действия фактора естественного отбора, о чем еще пойдет речь. Гены, контролирующие невоплощенные элементарные видовые и индивидуальные признаки, таким образом, могут быть без всякого вреда для особи и вида разрушены действием мутагенных факторов. Совокупность генов, контролирующих невоплощенные признаки, как с разрушенной, так и с цельной структурой должна образовывать своеобразный генетический балласт - третий блок информации в геноме.
Балластная часть ДНК может служить своеобразным внутренним щитом, берущим на себя часть разрушающего действия мутагенных факторов, и уменьшающим вероятность разрушения необходимой особи и виду наследственной информации.
В дополнение можно высказать утверждение, что так называемые "спящие" гены, о роли которых было высказано много суждений, никогда не "проснутся", так как отсутствует тот фундамент, на котором могли бы проявиться контролируемые ими признаки.
Кроме уже упоминавшихся совокупностей генов, образующих три блока генетической информации, геном любой биологической особи должен содержать еще один информационный блок - блок, обеспечивающий последовательность считывания генетической информации, используемой, в частности, во время развития эмбриона.
Необходимость существования такого блока ясна интуитивно, стоит только попытаться представить последовательность формирования эмбриона. Вопрос состоит в том, как этот блок информации реализован?
При работе блока считывания генетической информации должен безусловно соблюдаться следующий принцип: первым реализуется признак, который может быть реализован на более ранней стадии развития эмбриона - на меньшем количестве биологического материала. То же относится и к двум или более признакам, присутствующим в генетическом коде и имеющим сходный характер.
Ранее было сказано, что видоопределяющая совокупность генов моноинформативна, следовательно, случающиеся проявления у особей данного вида атавистических видовых признаков могут быть объяснены или воздействием на блок информации о видовой принадлежности родительской особи мутагенного фактора, или сбоями (под воздействием такого фактора) в работе блока считывания наследственной генетической информации на стадии развития эмбриона.
Сформулированный выше принцип работы блока считывания генетической информации имеет достаточно неожиданное следствие. Ранее было высказано утверждение, что процесс эволюции имеет однонаправленный характер - от простых видов к сложным. Представим теперь, что эволюционный путь некоего вида основан на нескольких блоках видооопределяющей информации, представляющих из себя соответствующие совокупности генов, которые в соответствии с порядком их внедрения в геном обозначим буквами A, B, C, D и E. Предположим, что очередной видообразующий вирус внедряет в геном вида блок видовой информации F, сходный по его содержанию, допустим, с блоком C. В том случае, если признаки, содержащиеся в блоке F могут быть реализованы на более ранней стадии развития эмбриона, чем признаки, содержащиеся в блоке C, они и будут реализованы, т.е. геном вида будет теперь состоять из блоков A, B, F, D и E. Но это в том случае, если признаки, содержащиеся в блоке F не уничтожают фундамент, на котором, пусть и частично, могут быть реализованы признаки, содержащиеся в блоках D и E. Рассматриваемая ситуация может привести к появлению вида близкородственного исходному. В том же случае, если после внедрения в геном блока F признаки, содержащиеся в блоках D и E не смогут быть реализованы, геном вида будет состоять из блоков видовой информации A, B и F, т.е. вид будет отброшен на более низкую ступень эволюции. Примеров иллюстрирующих такой ход событий я привести не могу, но возможность описанной ситуации в рамках новой теории не исключена, что позволяет объяснить некоторые зигзаги в видообразовании.
Глава 13
Естественный отбор и его место в новой теории
Наступил удобный момент поговорить о роли естественного отбора в существовании вида. Естественно, вопрос будет рассматриваться в самом общем плане воздействия естественного отбора на суммарный геном популяции.
Возьмем для рассмотрения факт преследования стаей волков лося по глубокому снегу. Исход такого преследования практически очевиден.
А теперь вспомним, как "опекает" волк стадо оленей в канадской тундре. Этот процесс великолепно описан Ф. Моуэтом в книге "Не кричи, волки!".
Формально между этими примерами разницы нет. И тут и там дело кончается гибелью особи не могущей отстоять свое право на жизнь. И оба эти примера могут служить примером действия фактора естественного отбора в соответствии с эволюционными теориями. Тем не менее, разница есть и она имеет принципиальный характер.
Неприспособленность лося к жизни в условиях глубокого снежного покрова имеет видовой характер. Но здесь имеет значение не этот факт, а тот, что особь выбраковывается по признаку, не имеющему отклонения от нормы. При этом, как ни странно это звучит в свете эволюционного подхода, естественный отбор не стимулирует появления нового вида, а сужает гамму индивидуальных признаков, которые погибшая особь могла бы передать потомству, обедняет генофонд популяции и вида в целом. В ситуации, описанной Ф. Моуэтом, особь отбраковывается потому, что она чаще всего либо больна, либо дефектна, во всяком случае, тот факт, что погибшая особь могла бы иметь полноценное потомство, далеко не очевиден. В этом случае отбраковка идет по факту наличия признака, имеющего отклонение от нормы, при этом фактор естественного отбора способствует санации генофонда, а значит, и продлению времени существования вида.
На основании приведенных примеров можно охарактеризовать одну из сторон естественного отбора.
Естественный отбор в устоявшемся биогеоценозе - это система взаимоотношений между видами, его образующими, позволяющая, в том числе, эффективно отбраковывать особи по критерию наличия у особи признака, имеющего отклонение от нормы.
Нетрудно заметить, что при таком определении процесс естественного отбора в устоявшемся биогеоценозе выступает в роли фактора, способствующего как можно более длительному существованию вида в неизменном виде, а если вернуться к моменту возникновения вида, то существованию вида в наиболее совершенной форме.
После рассмотрения роли естественного отбора в устоявшемся биогеоценозе логично рассмотреть к чему приводит действие фактора естественного отбора в неустановившемся биогеоценозе - при изменении условий обитания биологического вида или его популяции.
При изменении условий обитания естественный отбор по-прежнему проводит отбраковку особей по критерию наличия у них отклонений видовых и индивидуальных признаков от нормы, заданной при возникновении вида. Но параллельно - и это в рассматриваемом случае является определяющим - при изменении условий обитания в результате естественного отбора происходит отбраковка особей, у которых не проявлены признаки необходимые для выживания в изменившихся условиях. Это относится в первую очередь, а точнее - исключительно - к индивидуальным признакам, потому что видоопределяющая часть генома у всех особей, принадлежащих к данному виду, моноинформативна.
Вследствие существования рецессивных аллелей в блоке генетической информации об индивидуальных особенностях ни одна из особей не обладает всей гаммой индивидуальных признаков, присущих данному виду. Совокупностью генетической информации о всех индивидуальных признаках, которые могут быть проявлены у особей данного вида может обладать пусть и ограниченная по численности, но только определенная выборка особей.
А из этого напрямую следует, что в процессе естественного отбора обусловленного изменением условий обитания при отбраковке особей, не имеющих необходимых для выживания в изменившихся условиях признаков, происходит сокращение суммарной генетической информации, хранящейся в генофонде популяции и вида в целом об индивидуальных особенностях особей. Другими словами отбракованные особи уносят с собой генетическую информацию об индивидуальных признаках, которые могли бы оказаться полезными при очередном изменении условий обитания. Таким образом, в условиях неустановившегося биогеоценоза естественный отбор, способствуя выживанию популяции в меняющихся условиях, одновременно снижает приспособительный потенциал популяции.
Следствием из сказанного является то, что изолированная популяция при перманентном изменении условий обитания обречена на вымирание. С некоторыми оговорками это утверждение относится и к любому биологическому виду в целом.
Глава 14
Породы и псевдовиды. Попытка определения вида. Соотношение новой теории с эволюционными теориями видообразования
Предлагая по-новому взглянуть на проблему происхождения вида необходимо определиться как соотносится новая теория с уже имеющимися.
Есть ли возможность согласовать эти, казалось бы, взаимоисключающие друг друга теории? Попытаемся выяснить это на примере выведения все тех же новых собачьих пород.
Как происходит выведение новой породы? В качестве начальных условий предположим, что до настоящего момента селекционной работой с собаками никто не занимался. Наконец, два селекционера, каждый из которых отобрал из исходной популяции нужное количество производителей, приступают к такой работе - к воплощению, так сказать, своего видения собачьей породы.
Поскольку каждый селекционер оперирует не со всеми особями, составляющими вид, а с ограниченным числом производителей, то и число индивидуальных признаков, хранящихся в суммарном генофонде отобранных производителей меньше, чем таковых же в суммарном генофонде вида.
Чего добивается каждый селекционер? Чистоты полученной линии, т.е. отсутствия у потомства признаков, не вписывающихся в нарисованный каждым из них образ идеальной породы. При этом количество индивидуальных признаков в суммарном генофонде полученной в результате селекции линии или породы еще более уменьшается.
Вдобавок ко всему, новых признаков процесс селекции в геном не привносит. В итоге же выведенная линия, если представить ее как новую популяцию, обладает пониженной приспособляемостью к изменениям условий существования.
То же самое может произойти и происходит в природе в изолированных друг от друга популяциях, живущих в разных условиях, в результате естественного отбора.
Нелишним будет отметить, что в обоих случаях наличие определенных индивидуальных признаков становится определяющим фактором для выживания популяции. В обоих случаях в рамках популяции происходит увеличение процента мономорфности генов, контролирующих ставшие необходимыми для выживания индивидуальные признаки в полном соответствии с выводом, сделанным в третьей главе, что позволяет дать определение породы, возникающей в рамках биологического вида.
Порода - совокупность биологических особей, относящихся к определенному биологическому виду, для генома которых характерен повышенный в сравнении со средним для вида процент мономорфности составляющих его генов. При этом рост процента мономорфности происходит за счет генов, контролирующих индивидуальные признаки, присущие данной породе.
По причине того, что видение новой породы у каждого из селекционеров свое, сравнив результаты своей работы, они увидят между выведенными ими породами разительные отличия. В результате вполне может оказаться и так, что эти две породы окажутся репродуктивно изолированными, то есть при том, что особи, принадлежащие к каждой из выведенных пород не изменили своей видовой принадлежности, получение потомства при естественном способе скрещивания их с особями, принадлежащими к другим породам (равно как и с особями из исходной популяции), станет невозможным.
Опять же полные аналоги этому можно обнаружить и в природе - я имею в виду репродуктивно изолированные популяции.
Но поскольку ни селекционер, ни естественный отбор не могут сколь-нибудь заметно изменить видообразующую совокупность генов, видовые признаки продолжают оставаться одними и теми же (по отношению к представителям исходного вида) у представителей возникших пород, линий, репродуктивно изолированных популяций - как бы их ни назвали.
В результате полученные селекционерами или возникшие в природе под воздействием изменения условий обитания и естественного отбора линии, породы или популяции, достигшие стадии полной или частичной (кольцевые виды) репродуктивной изоляции, правильнее назвать псевдовидами. Процесс возникновения псевдовидов происходит с уменьшением суммарного количества индивидуальных признаков, хранимых в суммарном генофонде исходного вида или популяции и связанного с этим увеличением процента мономорфности генов, контролирующих индивидуальные признаки, т.е. сопровождается деградацией этого генофонда. Все теории, объясняющие возникновение псевдовидов, следует вследствие этого называть теориями деградационного псевдовидообразования.
Здесь уместно задать несколько вопросов. Почему, согласно эволюционным воззрениям, состояние репродуктивной изоляции популяции является необходимым предварительным условием возникновения нового вида? Ведь нового видового признака (признаков) у представителей таких популяций нет.
Что мешает формированию нового видового признака (признаков) в популяциях, не находящихся в состоянии репродуктивной изоляции? Что мешает такому формированию в том случае, если популяция изолирована географически?
Как будет происходить формирование нового видового признака (признаков) в популяции, достигшей состояния репродуктивной изоляции? Каким образом в результате такого формирования, если оно состоится, потеряют смысл выводы, сделанные в настоящей работе?
На мой взгляд, придание эволюционными теориями особого значения состоянию репродуктивной изолированности популяции вызвано простым заблуждением, основанном на том факте, что представители разных видов не могут скрещиваться между собой, как и представители репродуктивно изолированных популяций с представителями исходного вида.
Отсюда следует, что принципиальная ошибка эволюционных теорий состоит в подмене понятия "биологический вид" понятием "репродуктивно изолированная популяция".
В свете сказанного может быть предложена простая и логичная схема существования биологического вида.
Исходная популяция вида- предшественника. - Появление на ее базе нового вида при условии выполнения третьего постулата. - Разрастание популяции нового вида с освоением новых экологических ниш, дроблением на дочерние популяции и уже начинающимся процессом псевдовидообразования (из-за необходимости приспособления к отличающимся по условиям существования экологическим нишам. - Существование в освоенных экологических нишах с продолжающимся процессом псевдовидообразования из-за изменения условий существования внутри этих ниш. - Вымирание из-за исчерпания приспособительных возможностей вида, сопровождающего процесс псевдовидообразования и из-за накопившихся искажений наследственной генетической информации.
Сказанное может быть проиллюстрировано следующей схемой.
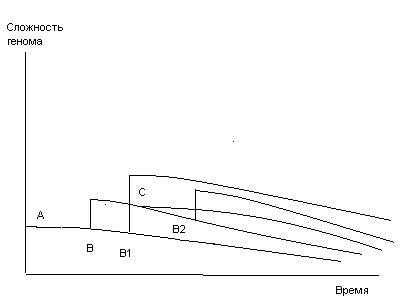
В отличие от традиционного изображения "эволюционного древа" или "куста" на данной схеме сложность генома вновь возникшего вида не возрастает со временем, а уменьшается, что обусловлено действием естественного отбора при изменении условий существования вида.
Точка А соответствует моменту возникновения некоего абстрактного исходного вида. Этим видом может быть и первая живая клетка. В этом случае расположение точки А на оси сложности генома отражает тот факт, что эта первая живая клетка должна была изначально обладать вполне определенной сложностью генома.
Точки В, В1, и В2 соответствуют моментам возникновения новых видов на основе предлагаемой теории. Точка С соответствует разделению вида на псевдовиды (нескрещивающиеся между собой популяции). Концы кривых, отображающих существование видов, соответствуют моментам их вымирания.
Таким образом, не подвергая сомнению ничего из сделанного в рамках существующих эволюционных теорий, я имею смелость утверждать, что они описывают не процессы, приводящие к возникновению новых видов, а процессы, сопровождающие существование и вымирание видов уже имеющихся.
Как нетрудно было заметить, предлагаемая теория видообразования утверждает, что состояние репродуктивной изоляции популяции является необходимым, но не достаточным критерием принадлежности составляющих ее особей к другому виду, также как и то, что способность к репродуктивному воспроизводству достаточный, но не необходимый критерий отнесения особей, взаимно способных к такому репродуктивному воспроизводству, к одному виду. Кроме того вообще способность к репродуктивному воспроизводству, так же как и состояние репродуктивной изолированности вторичны по отношению к генетической их обусловленности. Вследствие этого определение биологического вида должно строиться в первую очередь на генетической основе.
Но прежде чем попытаться сформулировать такое определение, необходимо еще раз оговорить, что в процессе формирования эмбриона и последующего развития особи реализуется далеко не вся генетическая информация, содержащаяся в геноме особи (вспомним, например о балластной части ДНК и о "спящих" генах). Учитывая этот момент, назовем ту часть генома, которая содержит генетическую информацию действительно используемую в процессе формирования и жизнедеятельности биологической особи реализуемой частью генома.
При этом условии, если оставить за рамками определения наличие половых различий или применять его к особям одного пола, определение вида может выглядеть следующим образом.
Биологический вид - совокупность биологических особей связанных идентичностью объема генов, составляющих реализуемую часть генома и соответствующего этой реализуемой части генома кариотипа. При этом аллели, составляющие реализуемую часть генома, у всех особей контролируют варианты одной и той же совокупности видовых и индивидуальных признаков.
Глава 15
Геном как физическая система
Утверждение, что генотип определяет фенотип, давно стало тривиальным и по
сути неоспоримым. Наглядное ему подтверждение - появление близнецов. В их
случае идентичный хромосомный набор и практически идентичные условия формирования
эмбриона приводят к появлению на свет двух или нескольких экземпляров фактически
одного и того же существа.
Но каким образом генотип формирует фенотип?
Если вспомнить еще не так давно действовавшее определение гена, согласно которому каждый конкретный ген отвечал за формирование конкретного признака (или участвовал в таком формировании), то единственное, что давало это определение, была иллюзия понимания сути дела.
Более современный подход, согласно которому назначение генов - это обеспечение производства белков и ферментов и вытекающий отсюда принцип: один ген - один белок (один фермент) или же один ген - одна полипептидная цепь - не оставляет даже места для существовавшей когда-то иллюзии.
Другими словами, геном уподоблен ныне заводу по производству стройматериалов, и при этом совершенно непонятно где же хранится чертеж в соответствии с которым используется произведенный продукт.
Результаты работ супругов Кирлиан, в частности известные фотографии древесного
листа с оборванным краем, на которых, тем не менее, контур изображения прорисовывается
таким образом, как если бы лист не был поврежден, дали основание говорить,
что "каркас", определяющий строение любого организма, формируется
"морфогенетическим" излучением, источником которого является сам
организм. При этом "морфогенетическое " излучение формирует "каркас"
организма по принципу голограммы.
Такому подходу, кстати, полностью соответствует эффект "фантомных болей"
- широко известный факт ощущения ампутированных конечностей и болей в них.
То есть после ампутации "каркас" ампутанта, формируемый излучением
нетронутых частей организма продолжает оставаться "частью" этого
организма, связанной с ним даже на уровне физических ощущений.
Теперь осталось только свести уже сказанное в этой главе воедино и предположить,
что источником этого самого "морфогенетического" излучения, формирующего
"каркас" или объемную голограмму любого живого организма является
геном каждой отдельно взятой клетки этого организма и их совокупности в целом.
Геном каждой отдельно взятой клетки, как и каждый участок голограммы, выполненной
традиционным способом, несет в себе информацию о строении всего организма,
то есть результатом "морфогенетического" излучения генома отдельной
клетки является "каркас" всего организма. И так же как в случае
традиционной голограммы, накопление в процессе эмбриогенеза количества клеток,
образующих растущий организм, суммирование их "морфогенетического"
излучения или, что то же самое, фрагментов голограммы, делает объемное "изображение
каркаса" все более детально "прорисованным".
Данное предположение придает геному клетки свойства не только биохимической,
но и физической системы. Какие же следствия выявляются в результате такого
подхода?
Во-первых. Становится крайне сомнительным наличие в геноме
любого организма "балластной части" ДНК, поскольку удаление любого
участка ДНК, даже не кодирующего полипептидные цепи, тем не менее изменяет
характеристики генома клетки как источника "морфогенетического"
излучения, что с необходимостью должно сказаться на "конструкции"
организма.
Во-вторых. К таким же последствиям и по тем же причинам должно приводить изменение
пространственного расположения хромосом в ядре клетки. Другими словами: изменение
пространственного расположения хромосом в ядре, если оно вследствие каких-либо
причин произойдет, с необходимостью должно приводить к появлению организма
на уровне генно-хромосомного состава неотличимого от исходного и тем не менее
представляющего из себя (если исходить из определения вида, приведенного ранее)
или новую породу, или псевдовид.
В-третьих. Можно предположить, что исходя из условия формирования четкой и
детальной картины "каркаса" организма и его отдельных органов, пространственное
расположение хромосом в клетках эмбриона до начала дифференциации клеток и
в клетках составляющих его органов после начала такой дифференциации и до
ее завершения должно быть взаимно согласованным.
В-четвертых. Если принять геном клетки за источник "морфогенетического"
излучения, то изменения аллельного состава генома (например, замена доминантного
аллеля рецессивным) должны, кроме всего, менять и параметры генома клетки
как "излучателя", а значит - отражаться на фенотипе особи. При таком
подходе можно снова вспомнить о той функции генов, которая приписывалась им
раньше - ответственность за формирование тех или иных признаков.
Признание за генно-хромосомным набором клетки функции физической
системы, то есть системы-источника "морфогенетического" излучения,
определяющего "конструкцию" живого организма, позволяет разделить
процессы "производства строительных материалов" и собственно "постройки"
организма.
Первый процесс при таком подходе обеспечивается собственно генами, кодирующими
полипептидные цепи, а второй - геномом клетки как единой пространственной
структурой.
Как следствие становится вполне понятным наличие большого числа идентичных
генов в геномах совершенно различных организмов - к примеру человека и дождевого
червя. Ведь и в строительстве громадное количество разнообразнейших зданий
возводится с использованием практически совпадающего набора стройматериалов
(кирпич, плиты перекрытий и так далее).
Таким образом, совпадения в номенклатуре генов, составляющих геном различных видов, практически ничего не говорят об эволюционном (в принятом смысле термина) происхождении одного вида от другого.
Тот факт, что процесс онтогенеза является непрерывным рядом изменений в том числе и в строении организма, заставляет предположить, что в процессе существования организма от рождения до смерти его геном с необходимостью также претерпевает непрерывные изменения, меняющие его характеристики как источника "морфогенетического" излучения. Можно предположить также (чисто из стремления не плодить излишние сущности), что механизм изменения генома как физической системы в процессе онтогенеза является единым как в плане обеспечения развития и достижения зрелости, так и в плане старения и, в конечном счете, смерти всего живого. А отсюда уже следует, что поняв механизм изменения строения генома как физической системы, принципиально возможно остановить процесс развития (старения) организма на любой его стадии.
Все приведенные в этой главе предположения могут быть на уже существующем уровне развития генной инженерии проверены экспериментально.
Нетрудно видеть, что признание за геномом свойств физической системы делает возникновение новых видов по схеме "мутации+отбор" еще более невероятным, чем это было показано ранее.
Заключение
Что устроено сложнее: атом водорода или атом урана? Ответ очевиден. Как появились атомы урана? В результате последовательного вовлечения атомов водорода и продуктов ядерного синтеза, начавшегося с атомов водорода, в процессы, происходящие в недрах звезд. Так, во всяком случае сейчас, отвечает на вопрос современная физика.
Налицо эволюция атома водорода в направлении к атому урана. Налицо инструмент этой эволюции - реакции ядерного синтеза.
Применительно к биологическим видам мы имеем в наличии процесс видообразования и можем воочию наблюдать его результаты. Предлагаемая теория видообразования предлагает в качестве возможного инструмента этой эволюции видообразующих вирусов. Что мы о них знаем?
А что мы знали полвека назад о реакциях ядерного синтеза?
Кроме того, вопрос о существовании видообразующих вирусов в конечном счете второстепенен. В основу предлагаемой теории в качестве краеугольного камня положен третий постулат, формулировку которого имеет смысл повторить.
Третий постулат: изменение информации о видовых и индивидуальных признаках в геноме вида-предшественника для возникновения нового видового признака (признаков) и, следовательно, для возникновения нового вида должно происходить внесением в геном единого блока информации, при взаимодействии которого с наследственной информацией, присущей виду-предшественнику, возникают все видовые и индивидуальные признаки нового вида.
Видообразующие вирусы в предлагаемой теории играют вспомогательную роль, как наиболее вероятный на настоящем уровне знаний фактор, способный претворить третий постулат в жизнь.
Тем не менее, я не могу избавиться от мысли, что все без исключения природные штаммы вирусов в свое время выполнили свою видообразующую функцию.
Томск - Барнаул
2002- 2007гг.